Haplogrupo H (ADNmt)
Em genética humana, o haplogrupo H (ADNmt) é um haplogrupo do ADN mitocondrial humano (ADNmt) tipico da Euroasia ocidental. Deriva do haplogrupo HV (ADNmt) e constitui o haplogrupo mais carateristico em todo o continente europeu, exceto entre os lapões. como pode ser observado seguindo a hiper-ligação.
{kind=link}
| Haplogrupo H | |
| Tempo de origem | 20,000-25,000 anos |
| Lugar de origem | SOAsia[1] |
| Ancestral | HV[1] |
| Descendentes | Linhagens H*, H1-39, 16129(H17+H27), 16129(H21+H30) |
|---|---|
| Mutações definidas | A2706A, C7028C[2] |
Origem e dispersão
Os calculos teóricos estimam uma origem provável no Médio Oriente[1] e posterior migração para a europa à 20.000 a 25.000 anos durante o pico da idade do gelo, provávelmente relacionada com a cultura gravetiana. Outros cálculos estimam uma idade de 15.000 a 20.000 anos[3] e desde que a maior diversidade encontra-se na região do Cáucaso, parte da Europa Oriental e do Médio Oriente seria a origem mais provável na Ásia Menor ou áreas adjacentes.
A descoberta de restos humanos conhecido como Paglicci 23 em Apúlia, Itália, datada de 28.000 anos atrás, e cuja análise genética revela pertencente ao haplogrupo HVR,[4] indica que a idade deste haplogrupo e sua migração pode ser maior.
No entanto, o haplogrupo H só foi encontrado na Europa, embora em baixa freqüência em restos humanos desde o início do Neolítico, à 7450 anos, três variantes de H1 e H23, H26, H46 e H88. A diversidade de haplogrupo H na Europa aparece a partir do Neolítico Médio em escombros por cerca de 6100-5500 anos, que também encontraram haplogrupos H3, H5, H7, H10, H16 e H89. Maior diversidade e aumentar a freqüência foi o resultado de contribuições genéticas substanciais de sucessivas culturas pan-europeias e, particularmente, a cultura do vaso campaniforme, que cresceram a partir da Península Ibérica no período neolítico tardio, 4.800 anos atrás. A partir de então espalhou-se H2, H3, H4, H11, H13, H16, além de H1.[5]
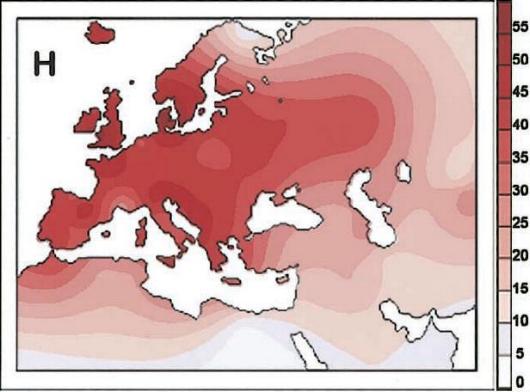
- Frequências do haplogrupo H1 no mundo (Ottoni et al. 2010)
| Região ou população | H1% | Nº de amostras |
|---|---|---|
| Africa | ||
| Tuaregues libios | 61 | 129 |
| Tuaregues (West Sahel) | 23.3 | 90 |
| Berbers (Morocco) | 20.2 | 217 |
| Morocco | 12.2 | 180 |
| Berbers (Tunisia) | 13.4 | 276 |
| Tunisia | 10.6 | 269 |
| Mozabite | 9.8 | 80 |
| Siwas (Egypt) | 1.1 | 184 |
| Western Sahara | 14.8 | 128 |
| Mauritania | 6.9 | 102 |
| Senegal | 0 | 100 |
| Fulani (Chad-Cameroon) | 0 | 186 |
| Cameroon | 0 | 142 |
| Chad | 0 | 77 |
| Buduma (Niger) | 0 | 30 |
| Nigeria | 0 | 69 |
| Ethiopia | 0 | 82 |
| Amhara (Ethiopia) | 0 | 90 |
| Oromo (Ethiopia) | 0 | 117 |
| Sierra Leone | 0 | 155 |
| Guineans (Guiné Bissau) | 0 | 372 |
| Mali | 0 | 83 |
| Kikuyu (Kenya) | 0 | 24 |
| Benin | 0 | 192 |
| Asia | ||
| Central Asia | 0.7 | 445 |
| Pakistan | 0 | 100 |
| Yakuts | 1.7 | 58 |
| Caucasus | ||
| Caucasus (north) | 8.8 | 68 |
| Caucasus (south) | 2.3 | 132 |
| Northwestern Caucasus | 4.7 | 234 |
| Armenians | 2.3 | 175 |
| Daghestan | 2.5 | 269 |
| Georgians | 1 | 193 |
| Karachay-Balkars | 4.4 | 203 |
| Ossetians | 2.4 | 296 |
| Europe | ||
| Andalusia | 24.3 | 103 |
| Basques (Spain) | 27.8 | 108 |
| Catalonia | 13.9 | 101 |
| Galicia | 17.7 | 266 |
| Pasiegos (Cantabria) | 23.5 | 51 |
| Portugal | 25.5 | 499 |
| Spain (miscellaneous) | 18.9 | 132 |
| Italy (north) | 11.5 | 322 |
| Italy (center) | 6.3 | 208 |
| Italy (south) | 8.7 | 206 |
| Sardinia | 17.9 | 106 |
| Sicily | 10 | 90 |
| Finland | 18 | 78 |
| Volga-Ural Finnic speakers | 13.6 | 125 |
| Basques (France) | 17.5 | 40 |
| Béarnaise | 14.8 | 27 |
| France | 12.3 | 106 |
| Estonia | 16.7 | 114 |
| Saami | 0 | 57 |
| Lithuania | 1.7 | 180 |
| Hungary | 11.3 | 303 |
| Czech Republic | 10.8 | 102 |
| Ukraine | 9.9 | 191 |
| Poland | 9.3 | 86 |
| Russia | 13.5 | 312 |
| Austria | 10.6 | 2487 |
| Germany | 6 | 100 |
| Romania | 9.4 | 360 |
| Netherlands | 8.8 | 34 |
| Greece (Aegean islands) | 1.6 | 247 |
| Greece (mainland) | 6.3 | 79 |
| Macedonia | 7.1 | 252 |
| Albania | 2.9 | 105 |
| Turks | 3.3 | 360 |
| Balcãs | 5.4 | 111 |
| Croácia | 8.3 | 84 |
| Slovaks | 7.6 | 119 |
| Eslovaquia (Oriental) | 16.8 | 137 |
| Eslovaquia (Ocidental) | 14.2 | 70 |
| Médio Oriente | ||
| Arabes Peninsula | 0 | 94 |
| Arabes Peninsula (incl. Yemen, Oman) | 0.8 | 493 |
| Druze | 3.4 | 58 |
| Dubai (United Arab Emirates) | 0.4 | 249 |
| Iraquianos | 1.9 | 206 |
| Jordanios | 1.7 | 173 |
| Lebaneses | 4.2 | 167 |
| Sirios | 0 | 159 |
| Eva mitocondrial (L) | ||||||||||||||||||||||||||||||||
| L0 | L1-6 | |||||||||||||||||||||||||||||||
| L1 | L2 | L3 | L4 | L5 | L6 | |||||||||||||||||||||||||||
| M | N | |||||||||||||||||||||||||||||||
| CZ | D | E | G | Q | A | S | R | I | W | X | Y | |||||||||||||||||||||
| C | Z | B | F | R0 | JT | P | U | |||||||||||||||||||||||||
| HV | J | T | K | |||||||||||||||||||||||||||||
| H | V | |||||||||||||||||||||||||||||||
Referências
- Achilli A, Rengo C, Magri C; et al. (2004). «The Molecular Dissection of mtDNA Haplogroup H Confirms That the Franco-Cantabrian Glacial Refuge Was a Major Source for the European Gene Pool». American Journal of Human Genetics. 75 (5): 910–8. PMC 1182122
 . PMID 15382008. doi:10.1086/425590
. PMID 15382008. doi:10.1086/425590 - van Oven M, Kayser M (2009). «Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation». Human Mutation. 30 (2): E386–94. PMID 18853457. doi:10.1002/humu.20921
- Pedro Soares et al 2009, Correcting for Purifying Selection: An Improved Human Mitochondrial Molecular Clock. y su página suplemento Arquivado em 29 de dezembro de 2009, no Wayback Machine. The American Journal of Human Genetics, Volume 84, Issue 6, 740-759, 04 June 2009
- D. Caramelli et al., A 28,000 Years Old Cro-Magnon mtDNA Sequence Differs from All Potentially Contaminating Modern Sequences. PLoS ONE, 2008
- Brotherton, Paul et.al. (2013) "Neolithic mitochondrial haplogroup H genomes and the genetic origins of Europeans" Nature Communications 4 (1764). doi:10.1038/ncomms2656